7. ПІСЛЯРАДІАЦІЙНЕ ВІДНОВЛЕННЯ РОСЛИН І ТВАРИН
2. Репараційне відновлення
В опроміненій клітині розрізняють два типи пошкодження – потенційно летальне та сублетальне. Потенційно летальними пошкодженнями називають такі, які можуть привести до загибелі клітини, але в певних умовах можуть бути відновлені. Під сублетальними розуміють такі типи пошкоджень, які самі по собі ще не приводять до загибелі клітини, але при наступному опроміненні здатні її викликати.
Відповідно розрізняють два типи репарації – від потенційно летальних і від сублетальних пошкоджень. Можливість репарації від сублетальних пошкоджень звичайно доводиться дослідами по фракціонованому опроміненню, в основі яких лежить гіпотеза про те, що якби радіаційне ураження носило повнієте незворотний характер, то ефект, викликаний фракціонованим опроміненням при певній сумарній дозі, був би таким же, як і при одноразовому опроміненні у тій же дозі. Але це не так. Величезна кількість даних, одержаних в дослідженнях з найрізноманітнішими організмами свідчить про те, що при фракціонуванні дози ступінь радіаційного ураження зменшується. При цьому ураження тим слабше, чим більший проміжок часу розділяє фракції і чим більша кількість фракцій, на які розділена доза.
В сухих системах (насінні, спорах, пилку), при обмеженні доступу кисню ефект фракціонування і потужності дози не проявляється або значно послаблюється. І найбільш переконливе пояснення цього явища можна знайти саме у визнанні реальності існування поклітинного відновлення, обов'язковою умовою якого є підтримування в клітинах активного обміну речовин.
Проте виявити природу відновлення за допомогою прийому фракціонування дози неможливо. Це вимагає проведення спеціальних досліджень з використанням сучасних методів молекулярної біології.
Молекулярне відновлення. Основним пошкодженням клітини при дії іонізуючих випромінювань, як уже не раз відзначалось, є пошкодження молекул ДНК. Головним структурним пошкодженням ДНК – одно- і двониткові розриви її полінуклеотидних ланцюгів. Репарація цих типів пошкоджень, яким приписується головна роль в загибелі клітини, вперше була показана в дослідах з бактеріями, які дають змогу одержувати штами, дефектні за окремими ферментами, що контролюють певні етапи репарації. На сл. 55 наведена схема основних етапів репарації однониткового розриву молекули ДНК.
Згідно неї, на першому етапі після утворення розриву відбувається виявлення місця пошкодження за допомогою спеціальних контролюючих систем. Потім ділянка полінуклеотидної нитки з пошкодженими нуклеотидами з обох боків надрізається за допомогою ферментів ендонуклеаз, вищеплюється з молекули – інцизія і видаляється – ексцизія. Останній етап здійснюється за допомогою ферментів екзонуклеаз. Розмір утвореного розриву буває різним – від декількох одиниць до декількох тисяч нуклеотидів. Вслід за цим на місці бреши відбувається комплементарний синтез ДНК з використанням залишку непошкодженої нитки ДНК в ролі матриці – так звана репаративна реплікація. Цей етап контролюється ферментами ДНК-полімеразами, які беруть участь і у звичайному реплікативному синтезі ДНК. І, нарешті, відбувається зшивання кінців синтезованої ділянки з полінуклеотидною ниткою.
Тип репарації, що йде за наведеною схемою, одержав назву ексцизійної репарації. Але описані й інші типи репараційного синтезу ДНК. Більшість з них хоч і має певні особливості, в цілому нагадують дану схему, суть якої визначається принципом "вищепління-заміщення".
Для репарації двониткових, або подвійних, розривів ДНК необхідно, щоб клітина мала активну систему рекомбінації і непошкоджені ділянки ДНК, гомологічні тим ділянкам, які мають подвійні розриви.
Існують дані і про молекулярне відновлення РНК, деяких білків, зокрема, ядерного білку хроматину, окремих основ ДНК. Але і в їх основі лежать механізми, аналогічні репарації ДНК. Тому репарація ДНК вважається основним механізмом молекулярного відновлення.
Відновлення клітинних структур. Ще в 1950-і роки була висунута гіпотеза про можливість відновлення радіаційних пошкоджень на рівні хромосом. Вона була заснована на підставі експериментальних даних про те, що при фракціонуванні дози опромінення кількість аберацій хромосом на клітину суттєво залежить від інтервалу часу між фракціями дози. Але якщо при невеликих інтервалах, вимірюваних хвилинами-годинами, практично завжди спостерігається зменшення числа аберацій, то при подальшому зростанні часу між фракціями або їх кількості результати виявляються досить суперечливими: в одних випадках відбувається їх зменшення до певного рівня, а в інших – після зменшення знову спостерігається збільшення. І практично всі дані, одержані як в дослідах з рослинами, так і з тваринами, описуються або кривою типу 1, або кривою типу 2, яка одержала назву кривої Лейна від імені автора, який вперше виявив таку залежність.
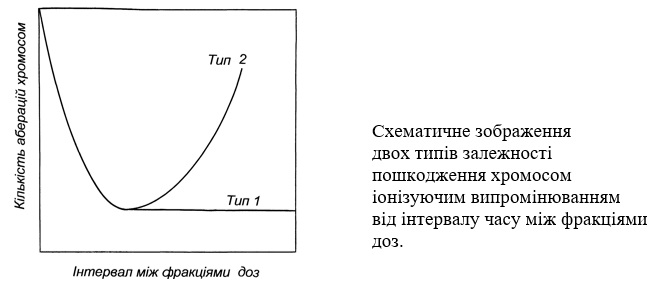
В цілому ж вважається, що в післярадіаційний період відбувається відновлення хромосом. Але досі ще не встановлено якихось певних механізмів їх репарації. Безперечно, що відновлення хромосом, що візуально спостерігається як зменшення кількості їх порушень, являє собою більш складний процес, ніж відновлення окремих молекул ДНК. Структурна організація хромосом еукаріотів досить складна. Крім ДНК до їх складу входить певна кількість РНК, різноманітні білки, з якими нуклеїнові кислоти утворюють міцні зв'язки. Важко уявити собі, щоб, наприклад, електрон міг зруйнувати таку складну структуру, а ще важче – механізм відновлення, який повинен охоплювати всі перераховані компоненти хромосом.
Є дані і про репараційне відновлення деяких інших структур клітини і, зокрема, мембран, які відіграють важливу роль в розвитку радіаційного ураження. Встановлено, що структура мембран, її проникливість, пошкоджені іонізуючими випромінюваннями, можуть з часом відновлюватися. Але молекулярні механізми цього процесу досліджені поки що слабо.
Яку роль відіграє репараційне відновлення в загальному відновленні багатоклітинного організму, невідомо. Досить переконливо не показана кореляція між радіостійкістю видів вищих організмів та їх здатністю до репарації. Відомі лише окремі роботи, в яких на основі дуже вузьких досліджень висловлюється версії про можливість такого зв'язку.
Безперечно, у збереженні кількості проліферуючих клітин процесам репарації повинно належати неабияке значення. Але розгляд часових характеристик кінетики відновлення будь-яких постійно оновлюваних тканин рослин і тварин свідчить про те, що для нормалізації здатності критичних органів до виконання своїх функцій необхідні, як мінімум, декілька клітинних циклів, тобто декілька діб, а не кілька годин, достатніх для поклітинного відновлення. І післярадіаційне відновлення будь-якої багатоклітинної системи є не стільки функцією репарації окремих її клітин, скільки розмноженням клітин, які зберегли здатність до поділу - репопуляції.
Шрифти
Розмір шрифта
Колір тексту
Колір тла
Кернінг шрифтів
Видимість картинок
Інтервал між літерами
Висота рядка
Виділити посилання
Вирівнювання тексту
Ширина абзацу