7. ПІСЛЯРАДІАЦІЙНЕ ВІДНОВЛЕННЯ РОСЛИН І ТВАРИН
| Сайт: | Навчально-інформаційний портал НУБіП України |
| Курс: | Сучасні проблеми радіобіології та радіоекології |
| Книга: | 7. ПІСЛЯРАДІАЦІЙНЕ ВІДНОВЛЕННЯ РОСЛИН І ТВАРИН |
| Надруковано: | Гість-користувач |
| Дата: | середа, 15 квітня 2026, 03:34 |
Опис
План лекції
1. Визначення радіобіології та її місце серед суміжних наук.
2. Напрями розвитку радіобіології та її задачі.
3. Історія радіобіології: етапи розвитку.
4. Сучасні проблеми радіобіології та радіоекології.
5. Необхідність широкої пропаганди радіобіологічних знань.
1. Післярадіаційне відновлення
Післярадіаційне відновлення організму – це відновлення після радіаційного ураження функцій клітин його критичних органів, що забезпечує нормалізацію їх функціональної діяльності і одужання його як цілісної системи. Таке визначення поняття "відновлення" з усією повнотою відбиває суть процесів, що приводять до нормалізації функцій багатоклітинного організму, пошкодженого іонізуючими випромінюваннями.
Проте цілком очевидно, що явище післярадіаційного відновлення являє собою багаторівневий процес, і те, що сприймається як відновлення організму, як одужання від променевої хвороби, звичайно відображає функціонування чисельних складних систем відновлення, які діють на різних рівнях організації.
Загальне відновлення досягається за рахунок чотирьох способів післярадіаційного відновлення: репараційного, репопуляційного, регенераційного і компенсаторного.
Репараційне, або поклітинне, відновлення, досягається за рахунок відновлення макромолекул клітини, в першу чергу ДНК, та окремих структур клітини – хромосом, мембран та інших. Репопуляційне відновлення забезпечується розмноженням клітин, які в момент опромінення перебували в радіостійкому стані і зберегли здатність до поділу. Регенераційне відновлення є результатом проліферації тканин і органів, що знаходяться в стані спокою. На відміну від інших організмів цей тип відновлення відіграє дуже важливу роль у вищих рослин. І, нарешті, компенсаторне відновлення, яке досягається за рахунок того, що функції пошкоджених клітин (тканин, органів), несучи підвищене навантаження, виконують неушкоджені клітини (тканини, органи), та за рахунок дедиференціації спеціалізованих клітин і тканин у проліферуючі.
2. Репараційне відновлення
В опроміненій клітині розрізняють два типи пошкодження – потенційно летальне та сублетальне. Потенційно летальними пошкодженнями називають такі, які можуть привести до загибелі клітини, але в певних умовах можуть бути відновлені. Під сублетальними розуміють такі типи пошкоджень, які самі по собі ще не приводять до загибелі клітини, але при наступному опроміненні здатні її викликати.
Відповідно розрізняють два типи репарації – від потенційно летальних і від сублетальних пошкоджень. Можливість репарації від сублетальних пошкоджень звичайно доводиться дослідами по фракціонованому опроміненню, в основі яких лежить гіпотеза про те, що якби радіаційне ураження носило повнієте незворотний характер, то ефект, викликаний фракціонованим опроміненням при певній сумарній дозі, був би таким же, як і при одноразовому опроміненні у тій же дозі. Але це не так. Величезна кількість даних, одержаних в дослідженнях з найрізноманітнішими організмами свідчить про те, що при фракціонуванні дози ступінь радіаційного ураження зменшується. При цьому ураження тим слабше, чим більший проміжок часу розділяє фракції і чим більша кількість фракцій, на які розділена доза.
В сухих системах (насінні, спорах, пилку), при обмеженні доступу кисню ефект фракціонування і потужності дози не проявляється або значно послаблюється. І найбільш переконливе пояснення цього явища можна знайти саме у визнанні реальності існування поклітинного відновлення, обов'язковою умовою якого є підтримування в клітинах активного обміну речовин.
Проте виявити природу відновлення за допомогою прийому фракціонування дози неможливо. Це вимагає проведення спеціальних досліджень з використанням сучасних методів молекулярної біології.
Молекулярне відновлення. Основним пошкодженням клітини при дії іонізуючих випромінювань, як уже не раз відзначалось, є пошкодження молекул ДНК. Головним структурним пошкодженням ДНК – одно- і двониткові розриви її полінуклеотидних ланцюгів. Репарація цих типів пошкоджень, яким приписується головна роль в загибелі клітини, вперше була показана в дослідах з бактеріями, які дають змогу одержувати штами, дефектні за окремими ферментами, що контролюють певні етапи репарації. На сл. 55 наведена схема основних етапів репарації однониткового розриву молекули ДНК.
Згідно неї, на першому етапі після утворення розриву відбувається виявлення місця пошкодження за допомогою спеціальних контролюючих систем. Потім ділянка полінуклеотидної нитки з пошкодженими нуклеотидами з обох боків надрізається за допомогою ферментів ендонуклеаз, вищеплюється з молекули – інцизія і видаляється – ексцизія. Останній етап здійснюється за допомогою ферментів екзонуклеаз. Розмір утвореного розриву буває різним – від декількох одиниць до декількох тисяч нуклеотидів. Вслід за цим на місці бреши відбувається комплементарний синтез ДНК з використанням залишку непошкодженої нитки ДНК в ролі матриці – так звана репаративна реплікація. Цей етап контролюється ферментами ДНК-полімеразами, які беруть участь і у звичайному реплікативному синтезі ДНК. І, нарешті, відбувається зшивання кінців синтезованої ділянки з полінуклеотидною ниткою.
Тип репарації, що йде за наведеною схемою, одержав назву ексцизійної репарації. Але описані й інші типи репараційного синтезу ДНК. Більшість з них хоч і має певні особливості, в цілому нагадують дану схему, суть якої визначається принципом "вищепління-заміщення".
Для репарації двониткових, або подвійних, розривів ДНК необхідно, щоб клітина мала активну систему рекомбінації і непошкоджені ділянки ДНК, гомологічні тим ділянкам, які мають подвійні розриви.
Існують дані і про молекулярне відновлення РНК, деяких білків, зокрема, ядерного білку хроматину, окремих основ ДНК. Але і в їх основі лежать механізми, аналогічні репарації ДНК. Тому репарація ДНК вважається основним механізмом молекулярного відновлення.
Відновлення клітинних структур. Ще в 1950-і роки була висунута гіпотеза про можливість відновлення радіаційних пошкоджень на рівні хромосом. Вона була заснована на підставі експериментальних даних про те, що при фракціонуванні дози опромінення кількість аберацій хромосом на клітину суттєво залежить від інтервалу часу між фракціями дози. Але якщо при невеликих інтервалах, вимірюваних хвилинами-годинами, практично завжди спостерігається зменшення числа аберацій, то при подальшому зростанні часу між фракціями або їх кількості результати виявляються досить суперечливими: в одних випадках відбувається їх зменшення до певного рівня, а в інших – після зменшення знову спостерігається збільшення. І практично всі дані, одержані як в дослідах з рослинами, так і з тваринами, описуються або кривою типу 1, або кривою типу 2, яка одержала назву кривої Лейна від імені автора, який вперше виявив таку залежність.
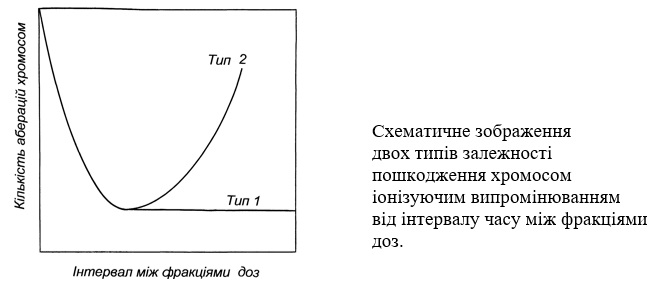
В цілому ж вважається, що в післярадіаційний період відбувається відновлення хромосом. Але досі ще не встановлено якихось певних механізмів їх репарації. Безперечно, що відновлення хромосом, що візуально спостерігається як зменшення кількості їх порушень, являє собою більш складний процес, ніж відновлення окремих молекул ДНК. Структурна організація хромосом еукаріотів досить складна. Крім ДНК до їх складу входить певна кількість РНК, різноманітні білки, з якими нуклеїнові кислоти утворюють міцні зв'язки. Важко уявити собі, щоб, наприклад, електрон міг зруйнувати таку складну структуру, а ще важче – механізм відновлення, який повинен охоплювати всі перераховані компоненти хромосом.
Є дані і про репараційне відновлення деяких інших структур клітини і, зокрема, мембран, які відіграють важливу роль в розвитку радіаційного ураження. Встановлено, що структура мембран, її проникливість, пошкоджені іонізуючими випромінюваннями, можуть з часом відновлюватися. Але молекулярні механізми цього процесу досліджені поки що слабо.
Яку роль відіграє репараційне відновлення в загальному відновленні багатоклітинного організму, невідомо. Досить переконливо не показана кореляція між радіостійкістю видів вищих організмів та їх здатністю до репарації. Відомі лише окремі роботи, в яких на основі дуже вузьких досліджень висловлюється версії про можливість такого зв'язку.
Безперечно, у збереженні кількості проліферуючих клітин процесам репарації повинно належати неабияке значення. Але розгляд часових характеристик кінетики відновлення будь-яких постійно оновлюваних тканин рослин і тварин свідчить про те, що для нормалізації здатності критичних органів до виконання своїх функцій необхідні, як мінімум, декілька клітинних циклів, тобто декілька діб, а не кілька годин, достатніх для поклітинного відновлення. І післярадіаційне відновлення будь-якої багатоклітинної системи є не стільки функцією репарації окремих її клітин, скільки розмноженням клітин, які зберегли здатність до поділу - репопуляції.
3. Репопуляційне відновлення
Джерелом репопуляційного відновлення є клітини, які зберегли репродуктивну здатність після опромінення. З одного боку, це клітини, що в момент дії радіації перебували в радіостійких періодах клітинного циклу, а з іншого боку – клітини, що знаходилися в стані спокою – "поза циклом".
Розмноження клітин, які перебувають в радіостійких фазах клітинного циклу. Радіостійкість клітин в залежності від фази і періоду клітинного циклу може розрізнятися в 2–3 рази. Тому завдяки асинхронності поділу, що є нічим іншим, як механізмом підтримки стійкості тканин до ушкоджуючих факторів, в популяціях твірних клітин в будь-який момент часу присутні клітини на різних фазах. І така клітинна популяція після опромінення являє собою досить строкату картину, так як в нiй в залежності від радіочутливості окремих клітин будуть присутні три основних їх пули: клітини, що втратили здатність до поділу (перебували в момент опромінення в найрадіочутливіших періодах циклу); клітини практично не пошкоджені (знаходились в найрадіостійкіших періодах), і клітини, що несуть потенціально летальні пошкодження, які в залежності від умов і ситуації можуть перейти як в клас 1, так і в клас 2 (які перебували в проміжних за радіочутливістю періодах клітинного циклу).
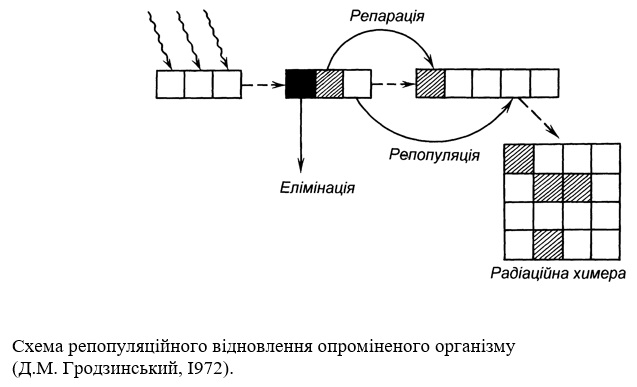
Клітини, які зберегли здатність до поділу, продовжують розмножуватися. Більше того, „відчувши” нестачу клітин в меристемі при елімінації тих, що втратили здатність до поділу, вони можуть ділитися з більшою швидкістю, ніж в нормі. Було показано, що тривалість клітинного циклу клітин меристеми кореня, котрі зберегли здатність до поділу, в такій ситуації скорочувалась з 17 до 1З годин – меристема прагне якнайшвидше відновити свій початковий клітинний об'єм, і після зменшення кількості клітин в меристемі незабаром після опромінення через деякий період, тривалість якого вимірюється кількома клітинними циклами, спостерігається її відновлення.
Розмноження клітин, які знаходяться поза циклом поділу. Резервом репопуляції є клітини в стані спокою – такі, що перебувають поза циклом поділу. У таких клітин є дві принципові можливості по відношенню до своєї подальшої долі: з припиненням дії чинника, який примусив їх вийте в стан спокою або після відновлення пошкодження, якщо зупинка була його наслідком, знову повернутися на шлях поділу, або перейти до диференціації. Яким саме з цих шляхів піде клітина, багато в чому визначається міжклітинними взаємодіями, які складаються в популяції.
Клітини в стані спокою мають більш високу радіостійкість, ніж ті, що діляться. І при дії випромінювань в дозах, що ведуть до втрати репродуктивної здатності у всіх або в певної кількості клітин, що діляться, до процесів репопуляці підключаються клітини, які перебувають поза циклом поділу, але які готові завжди приступити до поділу. Що служить сигналом для них? Безперечно, критична ситуація, що виникає в тканині, її спустошення. Саме тому перехід клітин в стан спокою варто розглядати як спосіб створення резервів тканини, які є фондом її репопуляційного відновлення.
Можливість репопуляційного відновлення і його темпи залежать від розміру пулу клітин, що зберегли здатність до поділу. Із збільшенням дози опромінення його об'єм зменшується і імовірність відновлення цим шляхом знижується. Теоретично репопуляційне відновлення можливе при наявності в популяції навіть однієї клітини, яка зберегла здатність до поділу. І дійсно, в дослідах з ссавцями було показано, що ті крипти кишкового епітелію, в яких виживає хоча б одна клітина, можуть повністю відновлюватися.
Слід відзначити, що найефективніший на теперішній час прийом лікування променевої хвороби людини, який полягає у трансплантації кісткового мозку здорового індивідууму хворому, являє собою ні що інше, як репопуляційне відновлення кровотворної тканини за рахунок розмноження непошкоджених радіацією клітин донора. Цей прийом застосовується саме тоді, коли при високих дозах опромінення втрачають повністю здатність до поділу стволові кровотворні клітини кісткового мозку.
4. Регенераційне відновлення
Якщо перші два шляхи післярадіаційного відновлення – репарація і репопуляція властиві всім багатоклітинним організмам, як рослинам, так і тваринам, то регенераційне відновлення – переважно лише рослинам. Еволюція прирекла більшість видів вищих рослин на прикріплений до постійного місця спосіб життя. I якщо тварини можуть укритися від несприятливих умов, які періодично виникають у навколишньому середовищі, то рослини такої можливості не мають. Саме тому поряд з такою надійною формою захисту виду, як існування надзвичайно стійкої фази їх онтогенезу – насіння, еволюція нагородила рослини потужною системою регенерації – здатності до відновлення втрачених органів і навіть всієї рослини за рахунок спеціальних тканин і органів, клітини яких, аналогічно клітинам насіння, знаходяться у стані спокою і мають високу стійкість до всіх уражуючих чинників.
Тут будуть розглянуті два основні типи регенерації у вищих рослин - регенерація тканин, які перебувають в стані спокою, і регенерація органів, які перебувають в стані спокою.
Регенерація тканин, які перебувають в стані спокою. На самому кінчику кореня, на межі з кореневим чохликом знаходиться особлива група клітин, яка має форму півсфери або двояко випуклої лінзи. Клітини цього утворення, яких нараховується не більше 1–2 тис., діляться дуже рідко – раз на 200–500 год., в той час як тривалість клітинного циклу переважної кількості оточуючих їх проліферуючих клітин меристеми складає лише 12–24 год. Англійський фізіолог рослин Ф. Клаус, який відкрив і перший почав дослідження цієї ділянки кореня у 1954 р., назвав його центром спокою.
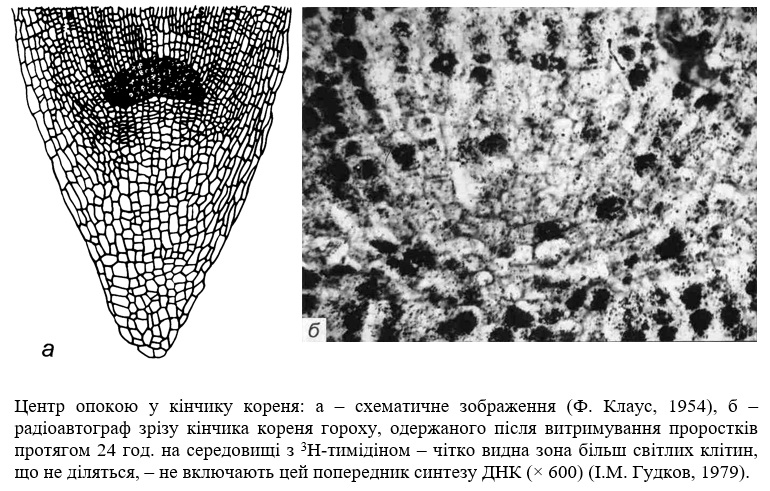
Функції центру спокою до кінця не з’ясовані. Саме тому деякі ботаніки, не вбачаючи його на анатомічних препаратах за звичайних методах фарбування клітин, взагалі ігнорують його існування. Але інші дослідники вважають його основою меристеми, з якої походять всі типи клітин кореня. Деякі дослідники вважають, що клітини центру спокою є надзвичайним резервом меристеми. При пошкодженні меристеми і втраті здатності до поділу її клітин клітини центру спокою починають ділитися і відновлюють первинний клітинний об'єм.
Існує певний критичний рівень пошкодження меристеми, при якому індукується поділ клітин центру спокою. Тільки при втраті здатності до поділу у певної кількості клітин до процесів репопуляційного відновлення можуть підключатися клітини центру спокою. Очевидно, це є результатом міжклітинних взаємодій, що існують між субпопуляціями клітин, які діляться і які перебувають в стані спокою.
Після повного відновлення меристеми, коли кількість клітин, що діляться, досягає в ній рівня норми, виникає новий центр спокою.
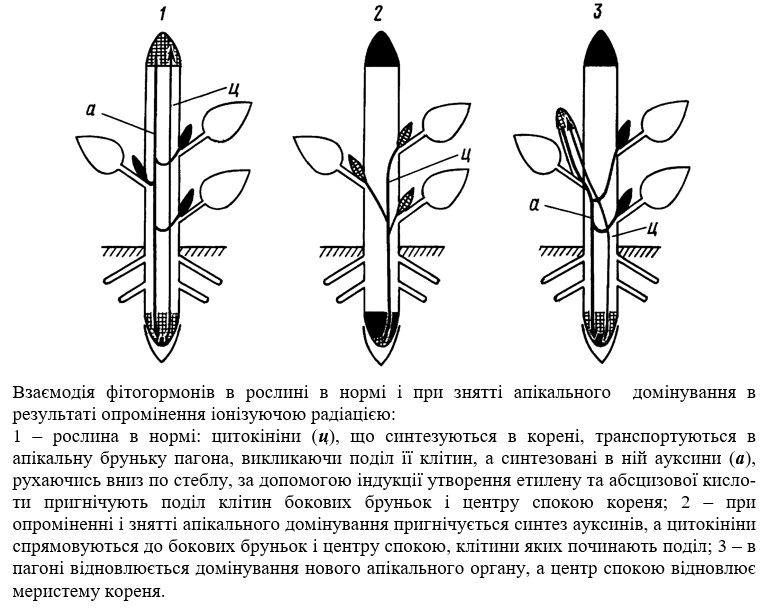
Регенерація органів, які перебувають в стані спокою. Верхівкова меристема пагона вищих рослин, яка активно росте, в значній мірі, а іноді і повністю, придушує поділ клітин в меристемах бокових бруньок. Це явище, що отримало назву апікального домінування, являє собою сильну міжклітинну взаємодію, яка відбувається на рівні цілісного організму. Видалення верхівки приводить до зняття апікального домінування і посилення проліферативної активності тканин, які перебувають в стані спокою і пробудженню бокових бруньок. Всі ці явища належать до регенераційних.
Апікальне домінування визначається гормональним типом впливу на точки росту. Це підтверджується чисельними дослідженнями, в яких за допомогою таких рістактивуючих речовин, як кінетин і гетероауксин, вдавалося знімати апікальне домінування або, навпаки, відновлювати його при видаленій верхівковій меристемі.
Опромінення рослин іонізуючою радіацією, природно, веде до більшого пошкодження верхівкових меристем, клітини яких активно діляться і ростуть. І якщо у рослини існують органи, які перебувають в стані спокою і мають підвищену радіостійкість, то її загибель повинна викликати їх регенерацію. І, безумовно, описані в радіобіологічній літературі факти, які свідчать про сильне гілкування опромінених рослин (дихотомія, фасціації), активацію утворення бокових коренів, про підвищення кущування та інше, пояснюється саме зняттям апікального домінування і посиленням регенераційних процесів.
Найкраще апікальне домінування вивчене у бобових рослин, які здавна вважаються класичними об'єктами для його демонстрації. Бокові бруньки у бобових розміщені в пазухах листків і тому називаються пазушними. За нормальних умов росту і розвитку вони знаходяться в стані спокою протягом всього онтогенезу. Але при знятті апікального домінування, наприклад, шляхом механічного видалення верхівкової бруньки, одна (іноді дві) бокова брунька вступає в ріст і розвивається в пагін, який повністю повторює організацію головного пагона і формує в подальшому всю рослину.
Аналогічна ситуація виникає при опроміненні рослин, як, до речі, і при пошкодженні апікальної бруньки хімічними агентами, її усиханні, відмерзанні і дії інших чинників. Існує мінімальне порогове значення дози опромінення, при якій апікальне домінування повністю знімається. Безумовно, воно залежить від видової радіочутливості рослини і визначається тим рівнем дози. який веде до припинення поділу у певної критичної кількості клітин і інгібування синтезу фітогормонів, що пригнічують ріст пазушних бруньок. Саме тоді клітини меристем пазушних бруньок приступають до проліферації.
Регенераційне відновлення дійсно слід віднести до найпотужнішої системи відновлення, якою еволюція нагородила вищі рослини, котрі не мають можливості захищатися від пошкоджень за допомогою інших засобів. З його основі лежать неспецифічні пристосувальні реакції, які розширюють межі зміни умов життя рослин, і механізм регенераційного відновлення ефективний при дії будь-якого пошкоджуючого агента.
5. Компенсаторне відновлення
Цей тип післярадіаційного відновлення є найменш вивченим в радіобіології, хоча компенсаторне посилення окремих функції у опромінених іонізуючими випромінюваннями організмів доводиться спостерігати нерідко. Серед них можна виділити два основних типи компенсаторних реакцій. Перший з них зв'язаний з виконанням непошкодженими внаслідок опромінення клітинами, тканинами, органами не властивих їм функцій. Другий – з посиленням функцій, коли непошкоджені клітини, тканини, органи посилюють власне навантаження з метою надолуження функції інших, які їх втратили.
Дедиференціація спеціалізованих клітин і тканин. У перебігу розвитку організму в ньому відбувається формування спеціалізованих клітин, тканин і органів за рахунок виникнення протягом морфогенезу відмінностей між однорідними клітинами і тканинами – процес диференціації. Так, меристематична клітина після здійснення декількох (п'яти-семи) поділів переходить до розтягнення, в процесі якого змінюється її форма і збільшуються розміри, потім до диференціації, при якій зовнішньо однакові клітини набувають різних властивостей та формують різні типи тканин, і далі до спеціалізації, яка характеризується різними рівнями метаболічної активності і ступенем структурної організації тканин і органів.
Звичайно, диференціація необоротна. Але в умовах пошкодження тканин, при злоякісному рості і деяких інших ситуаціях, що ведуть до зміни балансу фізіолого-активних речовин, природний хід руху клітин по життєвому циклу порушується, і вони можуть набути здатності до диференціації в зворотному напрямі, тобто дедиференціації. Саме такі умови можуть скластися при опроміненні рослин, коли, як згадувалося, змінюється баланс фітогормонів, і клітини меристеми втрачають здатність до поділу. При цьому диференційовані клітини починають ділитися і стають джерелом відновлення меристеми або виникнення нових твірних тканин.
Дедиференціація – один з найяскравіших прикладів компенсаторного відновлення, коли опромінені клітини набувають не властивих їм функцій.
До компенсаторного шляху післярадіаційного відновлення слід віднести нерідко констатоване після опроміненні посилення функцій клітин, тканин і органів, які зберегли свої функції. Так, при опроміненні кущових форм злаків при зменшенні кількості стебел ті, що вижили, як правило, бувають більш рослими і міцнішими, ніж окремі стебла в неопроміненому контролі. При пошкодженні генеративних органів і зменшенні кількості зав'язі на рослині сформовані колоски, стручки, плоди виявляються більш продуктивними, ніж в контролі. При зменшенні кількості зерен в суцвітті ті, що залишились, за абсолютною масою перевищують контрольні.
Ці факти можна пояснити з позицій зміни характеру перерозподілу поживних речовин, що надходять з кореневої системи – те, що призначалося для семи, дістається одному. В цьому відношенні не менш ілюстративними, але однозначно трактованими з позицій компенсаторного посилення функцій, є дані, одержані на рівні клітин, тканин. Зокрема, вище згадувалось, що при γ-опроміненні рослин в дозах, при яких кількість клітин в меристемах зменшується в 2–3 рази, різко зростає швидкість їх поділу. Виявляється, що клітини, які залишаються непошкодженими або відновили здатність до поділу, намагаючись поновити початковий клітинний об'єм меристеми, починають активно розмножуватися. В цей період після короткочасного гальмування швидкість поділу клітин суттєво зростала – майже на чверть скорочувалась тривалість їх клітинного циклу. Потім, по мірі досягнення меристемою норми, вона поступово збільшувалась і досягала початкового рівня.
Таке, навіть короткочасне, але досить суттєве посилення швидкості поділу клітин є характерним прикладом прояву компенсаторних явищ на рівні клітинної популяції.
Свідченням компенсаторного типу відновлення може бути нерідко спостережуване після опромінення у пошкоджуючих дозах тимчасове посилення синтезу нуклеїнових кислот, білків, фітогормонів, ферментів, макроергічних та інших сполук. Саме наслідком всіх цих процесів і є посилення поділу клітин, яке можна вважати їх інтегральним показником.
В цілому, розуміючі під терміном "відновлення” сукупність процесів, що забезпечують збереження організму від пошкодження або загибелі при дії іонізуючої радіації як системи, якій притаманне виконання певних функцій, варто уважно вивчати особливості прояву певних його реакцій, спрямованих на відновлення саме втрачених функцій. Управляючи цими процесами, можна сприяти більш або менш значному відновленню опромінених організмів у післярадіаційний період.
6. Управління процесами післярадіаційного відновлення
Буквально до 1960-х років була поширена думка, що наслідки радіаційного ураження не піддасться модифікації у післярадіаційний період. І хоча часто описувались зміни реакцій опромінених рослин і тварин протягом часу, їх залежність від різних фізичних факторів, це пояснювали звичайно дією на організм умов навколишнього середовища, які не мають безпосереднього відношення до процесів формування радіобіологічних ефектів.
Є не мало даних і про те, до багато хімічних речовин – ті ж відновники, солі різних металів і елементи живлення, гормони, ферменти, макроергічні сполуки та інші фізіологічно-активні речовини здатні суттєво впливати на хід розвитку радіаційного пошкодження при введенні їх в поживне середовище у післярадіаційний період.
Механізми дії більшості модифікаторів радіаційного ураження як фізичної, так і хімічної природи звичайно пов'язують з їх впливом не на віддалені етапи розвитку радіаційного ураження, а саме на первинні. Так, дія підвищених температур, вологості, газового стану, хімічних відновників пояснюється збільшенням швидкості рекомбінації вільних радикалів - фактором пошкодження, що реалізується в близькому післярадіаційному періоді. 3 відновлювальними ефектами на рівні репарації молекул ДНК і хромосом часто зв'язується дія низьких температур, ультрафіолетової радіації.
Дія деяких чинників хімічної природи, які застосовують перед опроміненням, може тривати і після нього, впливаючи на відновлювальні процеси. Так, безпосередньою участю в процесах післярадіаційного відновлення пояснюється протирадіаційна дія препаратів ДНК, окремих нуклеотидів, ферментів, гормонів та інших.
З інгібуванням процесів репараційного відновлення пов'язана радіосенсибілізуюча дія ряду хімічних речовин. Таку властивість, зокрема, мають специфічні інгібітори репарації ДНК кофеїн, акрифлавін, аміноптерін.
Ефективність репопуляційного та регенераційного відновлення визначається, головням чином, темпами розмноження клітин, які зберегли здатність до поділу після опромінення. Тому, діючи на швидкість їх поділу, можна управляти цими шляхами післярадіаційного відновлення. Так, створюючи оптимальні умови для вирощування рослин за допомогою регулювання температурного і газового режиму, освітленості, забезпечення елементами мінерального живлення, фітогормонами-активаторами росту можна сприяти ходу цих процесів. Саме це відкриває деякі, хоча і не такі вже й великі можливості до регуляції наслідків радіаційного ураження. Тому необхідно уважно вивчати і узагальнювати знання про влив різних факторів на наслідки радіаційного ураження організмів у відповідності зі шляхами післярадіаційного відновлення на різних рівнях організації живого.
Особлива стратегія повинна формуватися у післярадіаційний період щодо опромінених рослин. Як вже згадувалось вище, при аварійних ситуаціях на підприємствах ядерного паливного циклу і деяких інших гострому зовнішньому опроміненню і тривалому радіонуклідному забрудненню можуть бути піддані живі організми на досить значних територіях, що призводить до суттєвого радіаційного ураження біоценозу. Заходи, спрямовані на прискорення його післярадіаційного відновлення, повинні носити комплексний характер і по відношенню до рослинного компоненту визначатися двома основними прийомами: створенням для опромінених рослин в післярадіаційний період оптимальних умов для росту і розвитку та застосування фізіологічно-активних сполук, які прискорюють хід всіх типів післярадіаційного відновлення, а головним чином розмноження клітин, проліферацію тканин і органів, що зберегли ці функції.
Це може значно знизити наслідки радіаційного ураження фітоценозів, в тому числі і лісових угруповань, в умовах господарювання на забруднених радіонуклідами територіях, чому будуть присвячені наступні розділи.
7. Контрольні запитання до лекцій 4-7
1. Класифікація радіобіологічних ефектів.
2. Суть радіаційної стимуляції.
3. Основні типи морфологічних змін органів рослин при дії іонізуючих
випромінювань.
4. Вплив іонізуючих випромінювань на тривалість життя.
5. Специфіка радіаційної загибелі рослин.
6. Типи мутацій, що індукуються іонізуючими випромінюваннями.
7. Близькі та віддалені ефекти радіації.
8. Стохастичний характер віддалених наслідків радіаційного ураження.
9. Поняття про радіоміметики.
10. Відмінності між поняттями радіочутливість і радіостійкість.
11. Поняття ефективної дози. Півлетальні і летальні дози іонізуючих
випромінювань.
12. Найбільш радіочутливі і найбільш радіостійки види рослин.
13. Радіочутливість основних сільськогосподарських культур.
14. Порівняльна радіочутливість насіння і вегетуючих рослин.
15. Порівняльна радіочутливість видів тварин.
16. Специфіка дії іонізуючих випромінювань на фітоценози.
17. Причини широкої варіабельності радіочутливості видів.
18. Дія іонізуючих випромінювань на нуклеїнові кислоти.
19. Критичні органи рослин і тварин.
20. Особливості дії малих доз на живі організми.
21. Концепція безпороговості дози і поняття порогової дози – дві точки зору
в радіобіології.
22. Суть концепції ризику біологічної дії іонізуючої радіації.
23. Поняття модифікації радіаційного ураження організму.
24. Суть протирадіаційного біологічного захисту і сенсибілізації радіаційного
ураження.
25. Фізичні радіозахисні і радіосенсибілізуючі фактори.
26. Кисневий ефекту.
27. Фактор зміни дози (ФЗД) і його числові значення для радіозахисних і
радіосенсибілізуючих факторів.
28. Хімічні радіозахисні речовини.
29. Визначення поняття радіопротекторів та їх класифікація.
30. Радіоблокатори і радіодекорпоранти.
31. Визначення поняття післярадіаційного відновлення.
32. Класифікація типів післярадіаційного відновлення.
33. Види репараційного відновлення.
34. Механізм ексцизійної репарації ДНК.
35. Роль апікального домінування в регенераційному відновленні рослин.
36. Особливості компенсаторного відновлення.
Шрифти
Розмір шрифта
Колір тексту
Колір тла
Кернінг шрифтів
Видимість картинок
Інтервал між літерами
Висота рядка
Виділити посилання
Вирівнювання тексту
Ширина абзацу